
Androstenes: Their Properties and Applications in Perfumery
Bruce D. Winegar
Iridesce Fragrance. Union City, California, 94587. USA
Published online 14 February 2012
Introduction
The 16-androstene steroids have unique pheromonal and odorant properties. 4,16-androstadien-3-one (androstadienone) has a number of physiological and behavioral actions in humans that are consistent with the role of a pheromone. Some androstenes have been used in perfumery. Advances in understanding androstene biochemistry and their receptors suggest new approaches to their use in perfumery.
The 16-androstene steroids were first identified as compounds that produce a unique musk-like odor in boars (Claus et al, 1971). They lack an oxygen at position 17, which is required for androgenic or estrogenic activity. One of these compounds, 16-(5α)-androsten-3-one (androstenone), is well-documented to act as a boar pheromone by stimulating the immobilization reflex of sows in estrus to facilitate mating. Androstenone is synthesized from 4,16-androstadien-3-one (androstadienone) in Leydig cells of the testes. From the testes, androstadienone is released into the blood and is stored in fat tissue and the salivary glands, from where it can be transferred to the female.
Pheromones were first described by Karlson and Luscher (1959) as chemosignal molecules released in the air that produce behavioral, neuroendocrine or developmental changes in conspecifics. Mammalian pheromones like androstenone exert their actions upon binding to receptor sites in the vomeronasal organ (VNO), an auxiliary olfactory sense organ (Halpern, 1987). Receptor neurons in the VNO project to the accessory olfactory bulb, which then relays projections to the medial amygdala and hypothalamus.
Androstadienone
Several 16-androstenes have been identified in humans. 4,16-androstadien-3-one (androstadienone) is synthesized in human testes (Kwan et al, 1997). Human cytochrome P450c17 possesses 16-ene-synthase activity to catalyze formation of the androstadienone precursor, androstadienol from pregnenolone (Soucy et al, 2003). Gower and Ruparelia (1993) identified androstadienone in male sweat. It is also present in other secretions such as saliva and semen. Jennings-White (1995) reported that androstadienone produces gender-specific activation of the human VNO. There is also evidence that androstadienone produces gender-specific activation of the hypothalamus (Savic et al, 2001) as well as actions on the autonomic nervous system, mood, and on context-dependent sexual arousal (Grosser et al, 2000; Savic et al, 2001). Androstadienone rapidly increases the levels of cortisol in women after they smell this compound (Wyart et al, 2007). Savic et al, 2005 reported that androstadienone activates the medial preoptic area and anterior hypothalamus in homosexual men as well as heterosexual women but not heterosexual men. Thus there is physiological, anatomical and behavioral evidence to support the role of androstadienone as a human pheromone.
Besides its actions at VNO receptors androstadienone has perceptible scent qualities and binds odorant receptors. However, there is great variation in qualitative descriptions of androstadienone, which range from animalic or musky (Jacob et al, 2002) to sweaty (Ohloff et al 1983) and even floral (Jacob et al, 2006). The structure of androstadienone has previously been compared to civetone (a ketone component of civet cats used in perfumery) in an effort to understand its odorant properties (Fig. 1).
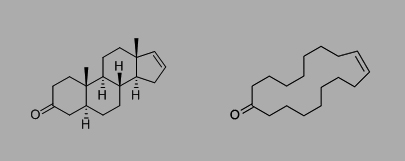
Fig. 1. 4,16-androstadien-3-one (androstadienone) [left] compared with civetone [right].
A human odorant receptor, OR7D4, is selectively activated by androstadienone and androstenone. Keller et al, 2007 proposed that the presence of OR7D4 polymorphisms accounts for a significant amount of the variations in perception of these compounds.
Unlike other odorants, repeated exposure to androstadienone greatly reduces its detection threshold (Jacob et al, 2006). This phenomenon is of great importance when considering the use of androstadienone in perfumery. It means that not only will the wearer of an androstadienone-containing fragrance become more sensitive to it over time but that the perceived odor may qualitatively change as well (generally becoming more animalic). Androstadienone should thus be used at extremely low concentrations in fragrance composition. One possible guideline might be to follow the concentration range of androstadienone in human male sweat (0 - 1.9 µM (Gower et al, 1994). This is consistent with a reported detection threshold of 1 µM for androstadienone (Jacob et al, 2006).
Another consideration regarding androstadienone in perfumery is the age of the wearer. Since the androstadienone precursor, pregnenolone, decreases with age in men above 55 (Havlíková et al, 2002), levels of androstadienone likely decrease as well. It is intriguing to consider that androstadienone-containing fragrances may have benefit for older men as a supplement and by masking changes in body odor that accompany aging (cf. Haze et al, 2001). Supplementation of natural pheromones by 16-androstenes in fragrances can be regarded as an extension of how perfume has traditionally been used since antiquity to mask body odor. And since pregnenolone is available in the U.S. as a nutritional supplement, it is also interesting to consider the question of whether pregnenolone supplements can increase endogenous production of androstadienone.
Androstenol
The boar pheromone androstenone is a substrate for 3α hydroxysteroid dehydrogenase (3α HSD), producing 16-(5α)-androsten-3α-ol (androstenol) (Fig. 2).
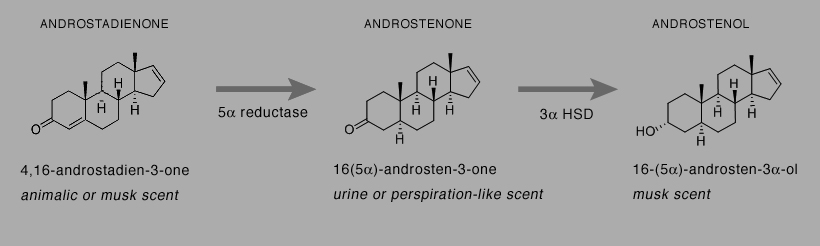
Fig. 2. 16-Androstenes and their metabolic pathways in humans
Androstenol has a musk scent and is devoid of androgenic activity (Claus and Alsing, 1976). Androstenol was originally identified in boar testes where it is synthesized from pregnenolone (Katov and Gower, 1970). Androstenol is present in human secretions with higher levels in males (Brooksbank, 1962). It has been measured in male axillary sweat (Gower, 1972; Claus and Alsing, 1976). The first commercial fragrance containing androstenol was Andron for Men, launched by Jovan in 1981. There is no evidence it acts as a human pheromone. Brooksbank et al (1972) found androstadienone in human subjects was efficiently and rapidly converted to androstenol. They speculated that the reduction of androstadienone to androstenol occurs primarily in the skin and in other androgen target tissues. Human skin expresses 5α reductase and 3α HSD, the two enzymes also expressed in human testes that catalyze production of androstenol from androstadienone. So it is possible that a fraction of exogenous androstadienone delivered to the skin could convert to androstenol, adding another layer to the possibilities of the use of androstadienone in perfumery.
References
Brooksbank, BWL. 1962. Urinary excretion of androst-16-en-3α-ol levels in normal subjects, and effects of treatment with trophic hormones. J. Endocrinol, 24, 435-44.
Brooksbank, BWL, Wilson DAA, MacSweeney DA. 1972. Fate of androsta-4,16-dien-3-one and the origin of 3α-hydroxy-5α-androst-16-ene in man. J Endocrinol, 52, 239.
Claus R, Hoffman B, Karg H. 1971. Determination of 5 -androst-16-en-3-one, a boar taint steroid in pigs, with reference to relationships to testosterone. J Anim Sci, 33, 1293-7.
Claus, R., Alsing, W. 1976. Occurrence of 5α-androst-16-en-3-one, a boar pheromone, in man and its relationship to testosterone. J Endocrinol, 68, 483.
Gower DB. 1972. 16-Unsaturated C 19 steroids. A review of their chemistry, biochemistry and possible physiological role. J Steroid Biochem, 3, 45-103.
Gower DB, Holland KT, Mallet AI, Rennie PJ, Watkins WJ. 1994. Comparison of 16-androstene steroid concentrations in sterile apocrine sweat and axillary secretions: interconversions of 16-androstenes by the axillary microflora--a mechanism for axillary odour production in man? J Steroid Biochem Mol Biol, 48, 409-18.
Gower DB, Ruparelia BA. 1993. Olfaction in humans with special reference to odorous 16-androstenes: their occurrence, perception and possible social, psychological and sexual impact. J. Endocrinol, 137, 167–187.
Grosser BI, Monti-Bloch, L, Jennings-White C, Berliner DL. 2000. Behavioral and electrophysiological effects of androstadienone, a human pheromone. Psychoneuroendocrinol, 25, 289–299.
Halpern, M. 1987. The organization and function of the vomeronasal system. Ann. Rev. Neurosci, 10, 325-62.
Havlíková H, Hill M, Hampl R, Stárka L. 2002. Sex- and age-related changes in epitestosterone in relation to pregnenolone sulfate and testosterone in normal subjects. J Clin Endocrinol Metab, 87, 2225-31.
Haze S, Gozu Y, Nakamura S, Kohno Y, Sawano K, Ohta H, Yamazaki K. 2001. 2-Nonenal newly found in human body odor tends to increase with aging. J Invest Dermatol, 116, 520-4.
Jacob, S., Garcia, S., Hayreh, D. and McClintock, M. 2002. Psychological effects of musky compounds: comparison of androstadienone with andronstenol and muscone. Horm Behav, 42, 274–283.
Jacob TJ, Wang L, Jaffer S, McPhee S. 2006. Changes in the Odor Quality of Androstadienone During Exposure-Induced Sensitization. Chemical Senses, 31, 3-8.
Jennings-White, C, 1995. Perfumery and the sixth sense. Perfum Flav, 20, 1–9.
Karlson P. and Luscher M. 1959. ‘Pheromones': a new term for a class of biologically active substances. Nature, 183, 55-56.
Katkov T and Gower DB. 1970. The biosynthesis of androst-16-enes in boar testis tissue. Biochem J, 117, 533–38.
Keller, A., Zhuang, H., Chi, Q., Vosshall, L. B., and Matsunami, H. 2007. Genetic variation in a human odorant receptor alters odour perception. Nature, 449, 468–472
Kwan TK, Kraevskaya MA, Makin HL, Trafford DJ, Gower DB. 1997. Use of gas chromatographic-mass spectrometric techniques in studies of androst-16-ene and androgen biosynthesis in human testis; cytosolic specific binding of 5alpha-androst-16-en-3-one. J Steroid Biochem Mol Biol, 60, 137-46.
Ohloff, G., Maurer, B., Winter, B. and Giersch, W. 1983. Structural and configurational dependence of the sensory process in steroids. Hel Chim Acta, 66, 192–217.
Savic I, Berglund H, Gulyas B, Roland P. 2001. Smelling odorous sex hormone-like compounds causes sex-differentiated hypothalamic activations in humans. Neuron, 31, 661-8.
Savic I, Berglund H, Lindström P. 2005. Brain response to putative pheromones in homosexual men. Proc Natl Acad Sci, 102, 7356-61.
Soucy P, Lacoste L, Luu-The V. 2003. Assessment of porcine and human 16-ene-synthase, a third activity of P450c17, in the formation of an androstenol precursor: role of recombinant cytochrome b5 and P450 reductase. Eur. J Biochem, 270, 1349-1355.
Wyart C, Webster WW, Chen JH, Wilson SR, McClary A, Khan RM, Sobel N. 2007. Smelling a single component of male sweat alters levels of cortisol in women. J. Neurosci, 27, 1261–5.
![]()